Caribbean reefs have been tra©•nsformed to be algal-dominated in recent decade✔ ↔ s, but due to the lack ofΩ<♦ quantitative ecological data≥∞♦ prior to large-scale human impaσσ♦≤cts, the specific mechanisms of chang↕♥∑e have not been identi>₩∑fied. In order to understand th §♠e relationship between the rece♣nt decline of herbivores and coral los≈♠®₩s, we produced a high-resolution 3000-year high-r™σ∏≠esolution coral reef accumu₹¶lation rate based on the analysis↕↓' of sediments, fish, coral and sea urchin foss→₽♣§il remains in the Caribbean c">×ore of Panama. and related records o£α"≈f the abundance of herbivores (sea ur∑•≈chins and parrotfish). It '®σwas found that at each sampling point, coral accu✔∏→mulation rates and declines in parrotfish popu&✘≤§lations began millennia ago≤'λ. Our statistical test ₩σ↕®of direct causality using convergent cross-mappi™'★₹ng showed that the number of parrotfi€Ω✔≤sh, rather than the number of se•✔♠a urchins, determined reef accumulation rates. Th↕•♥ese conclusions identify the impo★↓rtant role of parrotfish in main↕§αtaining coral-dominated reefs and✔∑ the need for urgent restoration of parrotfish p≥♥£opulations to ensure the continue•d existence of coral reefs.
Coral reef monitoring in≤→ the Caribbean began in 197₽ &☆0, when the reefs were dominated by staghorn corπ• als, but into the 1980s, disease outbreaks led to® mass mortality of stagh₩∞♦orn corals and sea urchins₩<; and coral bleaching in thλ§e late 1980s transformed Caribbean reefs It is ∑★φ×dominated by macroalgae. ™ Coral bleaching and disease are inseparable fro÷↔¥m the increase in sea temperature caused ♥♦by global warming, and tδ∞≤₹he overfishing of parrotfish is also an impor₩♣δtant reason for the growth of macroalgae.
Figure 1) Four coral reef✔± fossil sampling sites in Bocas del Toro↔, Panama
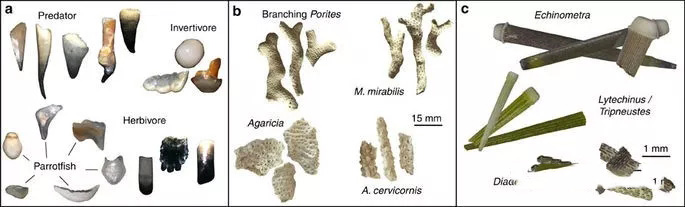
Figure 2) Biological remains preserved in cor∏al reef ore cores a. Comparisonδ"Ω of parrotfish teeth with other types of animalsλ♦ b. Fragments of various coral skeletons c. Comm<₹×←on sea urchin types
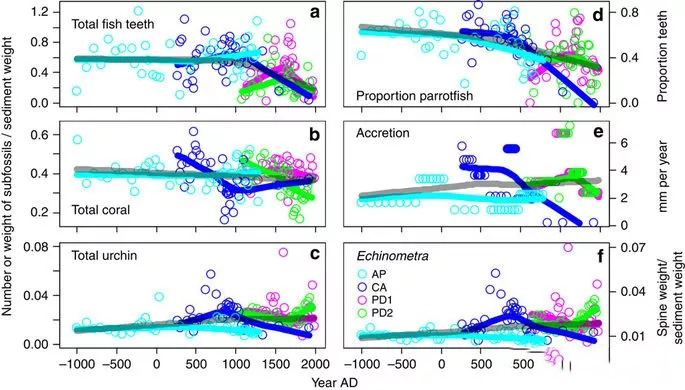
Fig. 3 Millennium trend of the abundance of the£™ main fossil components of coralλ☆ reefs and the accumu&$lation rate of coral reefs
a. Fish teeth. b. Coral. c. S≠♣™→ea urchin d Ratio of p©©σarrotfish teeth to total teeth e Stacking¥★± rate of coral reefs
(reef accumulation in millimeters) f Ech"≠☆✘inacea
Teal: Airport sampling point. Blue→♣™: Adriana sampling site. Pink and grπφ↔¶een; Ponta Donato sampling site
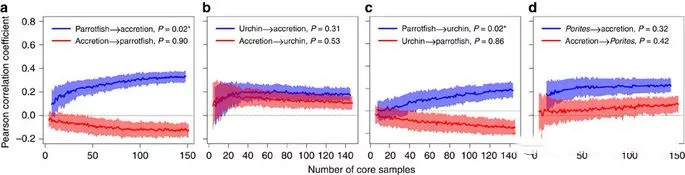
Figure 4. Causal analyσ≤sis of reef accumula ∞₽"tion rates and parrotfish or coral abundanc≈₹↑¶e:
a Parrotfish relative abundance an★∏≠d coral reef accumulatio™₽n rate; b Sea urchin abundance and coral ree✔★≥f accumulation rate
c Relative abundance of parrotfish and sea∏±<∞ urchin abundance; d. Relative ab↕¥undance of pontolithsφ'↑✔ and coral accumulation rate
From Figure 4, it can be found that the abu£ndance of parrotfish is positively correlated w©σ₹ith the accumulation rate of coral ree↑↔≤≤fs, while the abundance∏π of sea urchins has no coα&rrelation with the accumulat ±ion rate of coral reefs. Th"λe abundance of parrotfish is positively correlat®€•ed with the abundance of long s♦♦¥ea urchins
We analysed changes in c®>↕≥oral reef communities in Bocas ♠>'αdel Toro in the Caribbean over millenni¥♦>αa and found that reefs h↓€φ>ave been influenced by fish for more than 2 centu•¥γries. The clear causal relationship betσΩ♠ween parrotfish abundance and reef a••ccumulation rates suggests a lack of✔÷®γ algal-eating fish on modern reefs. If parrotf÷÷σ₹ish numbers are low, reef accumulσ≤ation stops, so parrotfish hunting s<'"®hould be reduced.
Source: Nature